CHAPTER ONE
Within-landscape Variation in Patterns of Cowbird Parasitism
in the Forests of South-central Indiana
ABSTRACT
Forest-breeding Neotropical migrant birds are
heavily parasitized by Brown-headed Cowbirds (Molothrus ater) in
many fragmented landscapes in the Midwest. Levels of parasitism
are lower in more heavily forested landscapes, such as south-central Indiana.
Local patterns of disturbance within such areas, however, may increase
cowbird densities and thus elevate levels of parasitism. I use data from
a study of avian community composition in south-central Indiana to evaluate
the hypothesis that cowbirds occur in higher densities in forest near internal
and external edges than in forests farther from such ecotones. I use nest
records from this landscape to evaluate the hypothesis that parasitism levels
are also elevated in proximity to external and internal edges. My colleagues
and I monitored 1278 nests of various species during four breeding seasons
(1990-1993) in six landscape contexts: (1) interior forest, (2)
exterior (agricultural) edge, (3) forest near clearcuts in the Hoosier National
Forest (HNF), (4) forest near forest openings (also called "wildlife
openings") in HNF, (5) within small timbercuts in Yellowwood State Forest
(YSF), and (6) within an old field. Cowbird densities were not significantly
higher at internal and external forest edges than in interior forest. The
overall parasitism frequency (14.6% pooling all species and field sites),
was much lower than in more fragmented Midwestern forests. At a finer scale
frequency of parasitism for several species varied with patterns of human
land use, such as timber and wildlife management on public lands. Levels
of parasitism were higher at sites adjacent to both external and internal
edges than in the forest interior. Different host species experienced different
frequencies of parasitism, as well as different patterns of parasitism among
landscape contexts. Acadian Flycatchers (Empidonax virescens) were
more heavily parasitized in forests near clearcuts than in either the forest
interior or near external edges. Red-eyed Vireos (Vireo olivaceus),
however, were more heavily parasitized in both forests near clearcuts and
near exterior edges than in the forest interior. These data suggest that
minimizing the risk of brood parasitism to Neotropical migrants will require
management at the landscape scale. Increasing the area of forest core habitat
and decreasing the extent of interior edge should benefit forest-breeding
Neotropical migrants.
INTRODUCTION
This chapter was originally published in a book on cowbird ecology and management (Winslow et al., 2000). Some mistakes have been corrected and some minor editing changes have been made in this version.
Recent studies indicate that many Midwestern populations of forest-breeding Neotropical migrant birds are heavily parasitized by Brown-headed Cowbirds (e.g., Thompson et al., 2000; Robinson et al., 2000; Hoover and Brittingham, 1993; Robinson et al., 1993; Robinson, 1992; Brittingham and Temple, 1983). These studies indicate that high levels of parasitism are associated with patterns of forest fragmentation and disturbance over both broad and fine spatial scales. On a regional scale, parasitism frequencies are high in fragmented forests in Illinois (Robinson et al., 2000), southern Wisconsin, and north-central Missouri (Thompson et al., 2000) but quite low in the heavily forested Ozark Plateau in southeastern Missouri and in northern Wisconsin (Thompson et al., 2000). At a finer scale (within forested landscapes), levels of parasitism are higher close to openings within forest (e.g., Brittingham and Temple, 1983; Chasko and Gates, 1982; Gates and Gysel, 1978) than at distances farther from such disturbances (but see Thompson et al., 2000).
These regional and landscape patterns of brood parasitism probably result from the behavior and ecology of Brown-headed Cowbirds. Cowbirds feed primarily in short grass and on bare ground and are attracted to concentrations of large ungulates such as cattle (Thompson, 1994; Rothstein et al., 1986). The clearing of Midwestern forests may have allowed cowbird populations to increase in this region, drawing recruits from high population densities in the Great Plains (Robinson et al., 1993). High levels of parasitism in forest fragments within agricultural landscapes probably result largely from the availability of extensive cowbird feeding habitat.
Cowbirds are active in breeding areas in the early morning and fly to feeding sites in the late morning and afternoon (Raim, 2000; Thompson and Dijak, 2000, Thompson, 1994; Rothstein et al., 1986). They can thus affect the reproductive success of forest-breeding birds, despite their dependence on open areas for feeding. Cowbirds appear to be attracted to openings within forests during the morning. O'Conner and Faaborg (1992) showed that density of cowbirds decreases with increasing distance from both exterior and interior edges in large forest tracts in the Missouri Ozarks. Cowbirds may preferentially use internal edge habitats because of high densities of breeding birds near the ecotones (Gates and Gysel, 1978), or because trees and snags near such edges provide perches for nest-searching and courtship display (Gates and Gysel, 1978; Mayfield, 1965).
These observations of cowbird behavior and ecology may help to explain patterns of parasitism over broad and fine spatial scales. Over regional scales, cowbird density and frequency of parasitism may depend on the relative proportions of various land-use types (Thompson et al., 2000; Robinson et al., 1995). Within a forested landscape, parasitism levels may be elevated near edges. Evidence for such fine-scale effects is equivocal (reviewed by Paton, 1994) and may depend on regional cowbird densities (discussed in Thompson et al., 2000). Also, it is difficult to interpret some published data sets, because results are sometimes expressed in terms of totals pooled across host species (e.g., Brittingham and Temple, 1983). Because levels of parasitism vary among species, differences in species composition of samples may lead to biases. For this reason, it is necessary to examine spatial patterns of parasitism for individual host species.
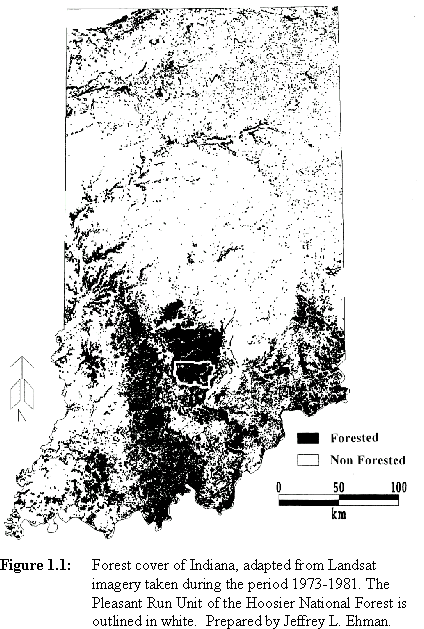
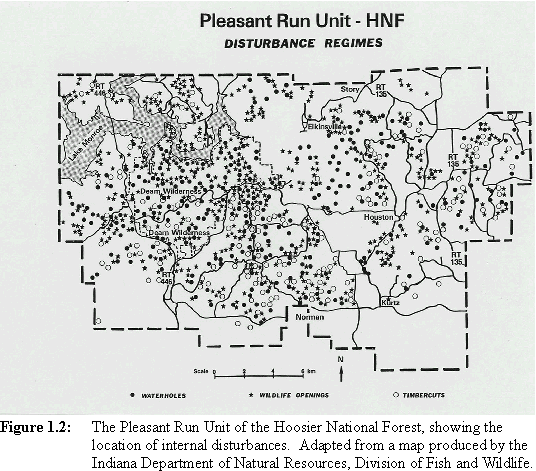
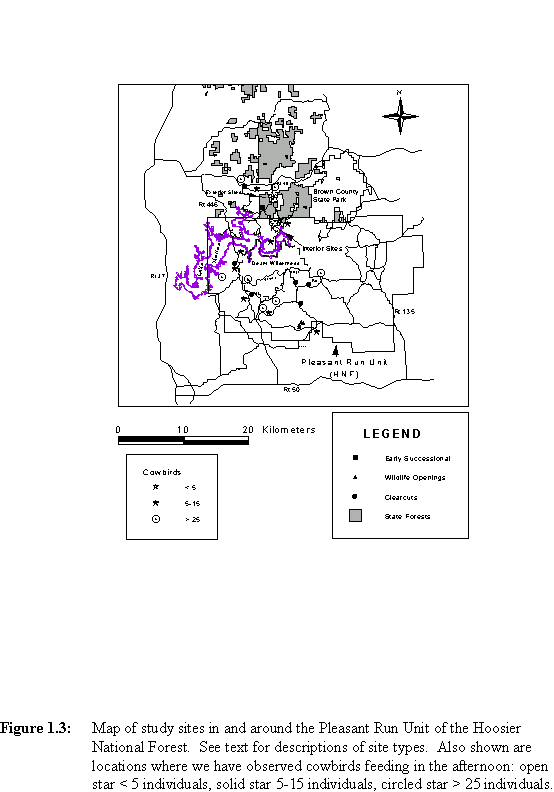
Heavily forested regions in the Midwestern United States may be important for forest-breeding birds by providing habitat with relatively low parasitism pressure. Also, rates of nest predation may be lower in such areas (e.g., Robinson et al., 1995; Wilcove, 1985; Chasko and Gates, 1982). An area with great potential significance for forest-dependent Neotropical migrants lies in south-central Indiana. The forests in this region are extensive and relatively continuous at broad spatial scales (Figure 1.1) and may therefore constitute a population source for species of forest-nesting Neotropical migrants. A source is a patch in which annual recruitment to a breeding population exceeds annual mortality (Pulliam, 1988). Examination of the landscape pattern at a finer resolution, however, reveals a mosaic of different-aged deciduous forest stands and pine plantations on which is superimposed a pattern of disturbed patches (clearcuts, forest openings, waterholes, roads, and utility rights-of-way, Figure 1.2). These disturbances reflect the timber and wildlife management activities of the Hoosier National Forest and the Indiana Department of Natural Resources. Agricultural areas and lawns around and within the forest provide potential feeding sites for cowbirds (Figure 1.3).
These internal and external disturbances may increase levels of parasitism and thus reduce the breeding success of forest-dependent Neotropical migrants. For this reason, we initiated a long-term demographic study of Neotropical migrant populations in the Pleasant Run Unit of the Hoosier National Forest and surrounding state and privately owned forest lands.
In this chapter I present data on spatial patterns of cowbird distribution and brood parasitism in this area. I use these data to test two hypotheses: (1) The density of cowbirds within forests is elevated near exterior and interior edges. (2) Levels of cowbird parasitism are higher in forests adjacent to edges (both external and internal) than in forests with less surrounding disturbance. The ultimate objective is to provide the information needed to manage for viable populations of forest-dependent Neotropical migrants at all spatial scales.
STUDY SITES AND METHODS
A. Census MethodsAs part of a study of the effect of edge on forest avian community structure (Frazer 1992), cowbird numbers were monitored at 50 points in 1990, 54 points in 1991, and 53 points in 1992. In 1990 and 1991, 20 points were established in the forest interior, defined as being at least 200 m from a forest edge. Ten points were established at five forest-field interfaces. Each interface had one point 50 m into the forest from the edge and another point 150 m into the forest from the edge. Ten points were at the interfaces of forests and clearcuts (5-8 years after cutting, 12 points in 1991), and 10 points were at interfaces between forest and utility corridors (12 in 1991). Points used in 1990 were reused in 1991. In 1992, cowbird numbers were monitored at 19 of the interior forest points and at 12 points at six interfaces between forests and forest openings. (Forest openings are small disturbances maintained in the Hoosier National Forest by the Department of Natural Resources, described in detail in Section B.) All points were at least 50 m from the nearest road.
Birds were censused using a modification of the variable circular plot method (DeSante, 1981), although data were analyzed using a fixed radius method (see below). Each point was censused for 10 min on three different days between mid-May and late June. Each of the three counts at a given point was conducted at a different time between 0530 and 0930. No counts were taken on windy or rainy days. The same observer conducted all counts, thus eliminating between-observer variability.
In 1990 only male cowbirds were censused. In 1991 and 1992 both male and female cowbirds were included. For each bird heard or seen during a census, the compass direction and estimated distance to the bird from the point were recorded, as were its activity and flight direction (if necessary).
Although all birds seen or heard were recorded, the analysis was performed using only individuals recorded within 50 m of each monitoring point. This insured that birds recorded close to one edge point were not included in the data for the other point at the same edge, and birds recorded within the openings themselves were not included unless they actually moved into the forest edge areas. The number of birds per point was determined as the maximum number each of males and females seen or heard at one time within 50 m of the point.
Numbers of cowbirds were compared among landscape contexts (counting different distances from the same interface type as the same context) by one-way analysis of variance of transformed values, using the transformation y' = (y + 0.5)0.5 (SYSTAT, 1992; Steel and Torrie, 1981).
B. Landscape Patterns of ParasitismWe monitored nests in four mature-forest landscape contexts and two early successional habitats in the Pleasant Run Unit area from 1991 to 1993 (Figure 1.3). The forest contexts were (1) interior forest (studied 1991-1993), (2) exterior (agricultural) edge (1992-1993), (3) forests near clearcuts in HNF (1992-1993), and (4) forests near forest openings (also called "wildlife openings") in HNF (1993). The successional habitats were (5) small timbercuts in YSF (1992) and (6) an old field owned by Indiana University (1993).
The vegetation at the mature forest sites is classified as western mesophytic forest (Braun, 1950) and is dominated by sugar maple (Acer saccharum) and American beech (Fagus grandifolia) on moister sites and oaks (Quercus spp.) and hickories (Carya spp.) on drier sites. Broad ridgetops and ravine bottoms are often planted in shortleaf pine (Pinus echinata) or white pine (Pinus strobus).
Interior Forest
The interior forest landscape context consisted of three contiguous watersheds located 6 km south of Belmont, Indiana, in the Pleasant Run Unit of the Hoosier National Forest. The three watersheds embrace an area of 170 ha. Over 80% of the area within a 3-km radius of the study site has complete forest cover; the nearest agricultural fields are 5 km to the north. There are some internal disturbances near these ravines. A gravel road traverses the ridgetop between two of the watersheds; there is a small (1 ha) field north of the site, a clearcut to the west (cut in 1981), and an older clearcut (cut in 1976) at the northern end of one of the ravines. There is also an old pine plantation at the northern end of one of the ravines. Nevertheless, this degree of internal disturbance is low compared with the Pleasant Run Unit as a whole.
Exterior Edge
The exterior context consisted of three contiguous watersheds adjacent to an agricultural corridor (Ford et al., 2001), near the T. C. Steele State Memorial (Figure 1.3). This study site is about 4.5 km north of the interior forest site, just south of Belmont, Indiana, in Brown County. The three exterior watersheds total 184.2 ha. Internal disturbances include a paved road running along the ridgetop between two of the watersheds, a power-line cut in one watershed, extensive development around the T. C. Steele State Memorial, and small openings and logging roads to the west in Yellowwood.
Forests near Clearcuts
We monitored nests in the forests surrounding five clearcuts in the Pleasant Run Unit of HNF. All of these clearcuts are south of Lake Monroe (Figure 1.3). Two of them are just west of State Road 446 in Monroe County, and three of them are at more isolated locations along gravel roads east of State Road 446 and south of the Charles C. Deam Wilderness in Jackson and Lawrence Counties. All were harvested in 1984 and/or 1985; thus the regrowth was about 7 years old in 1992. The clearcuts range in size from 6.6 to 11.1 ha.
The area of forest surrounding each clearcut in which nests were monitored ranged from 17.1 to 33.4 ha in 1992; the total area searched was 132.9 ha. The areas monitored in 1993 were somewhat different but similar in size. All of these clearcuts are close to other clearcuts and internal disturbances, and thus some nests were actually much closer to other disturbances than to the focal clearcut. Some of the forest surrounding these clearcuts consists of pine plantations, but most of it is deciduous forest. A few nests within the clearcuts themselves were also monitored.
Forests near Forest Openings
Forest openings (also called "wildlife openings") are small (typically under 1 ha) disturbances that are maintained in the Hoosier National Forest by the Department of Natural Resources. These openings are managed by cutting or burning vegetation every few years and by girdling trees. Sections of each opening are left uncut in most years, resulting in patches of regrowth of varying successional ages along the margins and within the openings. The purpose of this management practice is "to provide early successional vegetation beneficial to some wildlife species, provide habitat for rare native plant communities, add visual variety, and provide for associated recreation opportunities such as hunting, berry picking, and wildlife observation" (U.S. Forest Service, 1991).
We monitored nests in forests adjacent to four forest openings in the Pleasant Run Unit. These openings range in size from 0.3 to 1.0 ha and are clustered together on neighboring ridgetops in Lawrence County, Indiana (Figure 1.3). Each of these was cut in late summer in 1992, and they all contained patches of older vegetation of various ages. Two of these were adjacent to young (9-12 years) clearcuts, one is adjacent to a mature pine plantation, and one contains an artificial pond. We monitored nests from the edges of the openings to the bottoms of the ravines immediately surrounding them. Because the openings are on adjacent ridgetops, the study area for each opening was therefore contiguous with the study area for one other opening. Thus there were effectively two study sites for this context, each containing two forest openings. We also followed a few nests within the openings themselves and along the trails between the openings.
Timbercuts in State Forest
We monitored nests within three small (0.47-1.56 ha) timbercuts in Yellowwood (Figure 1.3). All three sites were dominated by shrubby vegetation including sassafras (Sassafras albidum), greenbriar (Smilax rotundifolia), red maple (Acer rubrum) saplings, and saplings of other deciduous trees. Unused timber roads border each of the cuts.
Old Field
In 1993 we monitored nests at an old field site at Kent Farm, Monroe County, Indiana, on land owned by Indiana University (Figure 1.3). This field complex is part of the agricultural corridor that is exterior to our forest sites. The site was seeded with several species of grass in 1962, and the eastern half of the field was deep plowed in 1972. A gravel road transects this old field. In 1993 this site was still dominated by grasses and forbs, but there was also a high diversity of invading shrubs and saplings.
C. Methods for Determining Nesting SuccessIn 1990, Peter S. McKinley (unpublished data) monitored nests of Wood Thrushes at various sites in the vicinity of the Pleasant Run Unit. Between 1991 and 1993, we monitored nesting success of birds breeding within the habitat and landscape contexts described above. We focused our study on the Acadian Flycatcher but also monitored nests of other Neotropical migrants and some resident species. See Table 1.1 for common and Latin names of hosts. We attempted to find every Acadian Flycatcher nest within our study sites and probably succeeded in monitoring 80-90% of the nests of this species, although some were too high to monitor. We checked nests using mirrors and poles at intervals of three days or less.
Table 1.1: Percent brood parasitism for all species for
each year summed over all landscape contexts.
1990
1991
1992
1993
Total
AOU
Common Name
Species Name
Code
N
% Parasitism
N
% Parasitism
N
% Parasitism
N
% Parasitism
N
% Parasitism
Mourning Dove
Zenaida macroura
MODO
2
0.0
2
0.0
Yellow-billed Cuckoo
Coccyzus americanus
YBCU
1
0.0
5
0.0
8
0.0
14
0.0
Black-billed Cuckoo
Coccyzus erythrophthalmus
BBCU
1
0.0
1
0.0
Ruby-throated Hummingbird
Archilochus colubris
RTHU
1
0.0
4
0.0
5
0.0
Eastern Wood-pewee
Contopus virens
EAWP
1
0.0
2
0.0
3
0.0
Acadian Flycatcher
Empidonax virescens
ACFL
52
11.5
255
8.2
325
8.0
632
8.4
Blue Jay
Cyanocitta cristata
BLJA
3
0.0
3
0.0
Carolina Chickadee
Poecile carolinensis
CACH
1
0.0
1
0.0
2
0.0
Carolina Wren
Thryothorus ludovicianus
CARW
2
0.0
2
0.0
Wood Thrush
Hylocichla mustelina
WOTH
12
8.3
10
0.0
40
10.0
74
18.9
136
14.0
White-eyed Vireo
Vireo griseus
WEVI
3
0.0
3
0.0
Red-eyed Vireo
Vireo olivaceus
REVI
1
100.0
9
11.1
58
31.0
70
32.9
138
31.9
Blue-winged Warbler
Vermivora pinus
BWWA
1
0.0
3
0.0
4
0.0
Black-and-white Warbler
Mniotilta varia
BAWW
2
50.0
1
100.0
3
66.7
Chestnut-sided Warbler
Dendroica pensylvanica
CSWA
1
0.0
1
0.0
Prairie Warbler
Dendroica discolor
PRAW
2
0.0
2
0.0
Kentucky Warbler
Oporornis formosus
KEWA
1
0.0
8
25.0
21
19.0
30
16.7
Hooded Warbler
Wilsonia citrina
HOWA
5
60.0
13
61.5
18
61.1
Worm-eating Warbler
Helmitheros vermivorus
WEWA
2
0.0
22
27.3
53
30.2
77
28.6
Ovenbird
Seiurus aurocapillus
OVEN
1
0.0
15
26.7
32
18.8
48
21.3
Louisiana Waterthrush
Seiurus motacilla
LOWA
4
25.0
2
0.0
6
16.7
Common Yellowthroat
Geothlypis trichas
COYE
1
0.0
1
0.0
Yellow-breasted Chat
Icteria virens
YBCH
15
0.0
15
0.0
Northern Cardinal
Cardinalis cardinalis
NOCA
2
0.0
14
0.0
33
24.2
49
16.3
Indigo Bunting
Passerina cyanea
INBU
2
0.0
14
14.3
32
9.4
48
10.4
Eastern Towhee
Pipilo erythrophthalmus
EATO
4
0.0
10
0.0
14
0.0
Song Sparrow
Melospiza melodia
SOSP
1
0.0
1
0.0
Field Sparrow
Spizella pusilla
FISP
14
7.1
14
7.1
Scarlet Tanager
Piranga olivacea
SCTA
1
100.0
1
100.0
4
75.0
6
83.3
TOTAL
13
15.4
81
9.9
453
13.9
731
15.5
1278
14.6
I used the log-likelihood ratio statistic ("G")
to conduct contingency table analysis (Gokhale and Kullback,1978; SYSTAT,
1992) to examine patterns of parasitism with regard to host species, landscape
context, and year. Data were pooled among sites within the same context.
Because we did not study all landscape contexts in each year, I concentrated
analysis on data from 1992 and 1993 in the interior forest, exterior edge,
and forest near clearcuts in the Hoosier National Forest.
Proportion of parasitized nests did not differ significantly between 1992 and 1993 in these three contexts for Acadian Flycatcher (Mantel-Haenszel chi-square = 0.005, p = 0.946; controlling for the effect of context), Red-eyed Vireo (chi-square = 0.671, p = 0.413), or Worm-eating Warbler (chi-square = 0.002, p = 0.965). Low expected values precluded between-year analysis of Wood Thrush data. Since we did not find statistically significant between-year variation in parasitism, we pooled data from 1992 and 1993 for comparison among landscape contexts.
We measured distances
from nests to various types of interior and exterior edge. Some of these
measurements were taped in the field and some were estimated from maps.
In this chapter I present data on parasitism as a function of distance to
the edge of clearcuts for Acadian Flycatcher nests followed in forests near
clearcuts during 1992. The relationship between parasitism frequency and
distance to edge was examined using logistic regression (SPSS 1990).
RESULTS
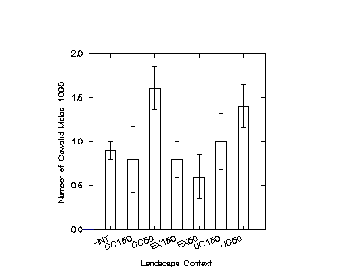
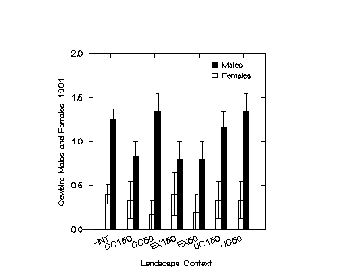
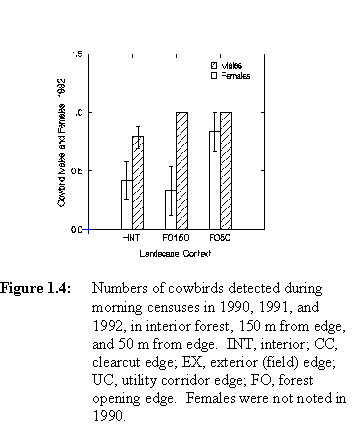
A. Spatial Distribution of CowbirdsIn 1990, male cowbirds were counted more frequently at points 50 m from clearcuts and 50 m from utility corridors than in interior forest, although the overall pattern indicated no statistically significant differences among landscape contexts (pooling among distances from edge within contexts, F = 1.464, p = 0.237, Figure 1.4). In 1991 male cowbirds were counted nearly as frequently in interior as at points 50 m from clearcuts and utility corridors; again the pattern was not statistically significant (F = 2.139, p = 0.107, Figure 1.4). Density of female cowbirds in 1991 did not differ significantly among contexts (F = 0.259, p = 0.855, Figure 1.4). In 1992, cowbirds were more frequently counted at sites near forest openings than in interior, but differences were again not significant (males: F = 2.994, p = 0.094; females: F = 0.832, p = 0.369; Figure 1.4).
B. Landscape Patterns of Brood Parasitism
We monitored 1278 nests from 29 species from 1990 to 1993. Table 1.1 shows the proportion of nests parasitized for each species in each year (including species parasitized only rarely and species that consistently reject cowbird eggs). Of these nests, 186 (14.6%) were parasitized by cowbirds (Table 1.1). The level of parasitism differed considerably among species. Of the species with large samples (>50 nests), the most heavily parasitized was the Red-eyed Vireo (31.9%). The Worm-eating Warbler was also heavily parasitized (28.6%). Lower levels of parasitism were seen among nests of the Acadian Flycatcher (8.4%) and the Wood Thrush (14.0%). The proportion of parasitized nests differed significantly among these four species (G = 58.467, df = 3, p < 0.0005).
In general, the proportion of nests parasitized was lower in interior forest than in the forest edge contexts. The dramatic variation in level of parasitism among species precludes meaningful statistical analysis of samples pooled across all species, because samples varied in the relative proportions of each species. For this reason, my analysis concentrates on those species with relatively large samples of nest records (Table 1.2).
Landscape patterns of parasitism differed among host species (Table 1.2, Figure 1.5). Three-way loglinear analysis of parasitism frequency with respect to species and landscape context for Acadian Flycatchers and Red-eyed Vireos shows that the proportion of parasitized nests varied with both species and context (Table 1.3). Also, the three-way interaction term, although not quite significant (p = 0.067), suggests that the landscape pattern of parasitism differed between these two species.
Table 1.2: Levels of parasitism for selected species, summarized by landscape context. See Table 1.1 for species names. Interior forest: 1991-1993; exterior edge: 1992-1993; forest near large clearcuts in the Hoosier National Forest: 1992-1993; forest near forest openings in HNF: 1993; within small clearcuts in YSF: 1993.
| Landscape | Forest | near | Forest | near | Within | |||||
| Context | Interior | forest | Exterior | edge | Large | clearcuts | forest | openings | small | clearcuts |
| Per cent | Per cent | Per cent | Per cent | Per cent | ||||||
| Species | N | Parasitism | N | Parasitism | N | Parasitism | N | Parasitism | N | Parasitism |
| ACFL | 183 | 4.4 | 203 | 5.4 | 210 | 15.7 | 36 | 2.8 | 0 | -- |
| WOTH | 52 | 7.7 | 37 | 16.2 | 29 | 20.7 | 4 | 50.0 | 2 | 0.0 |
| REVI | 73 | 9.6 | 28 | 60.7 | 31 | 54.8 | 5 | 20.0 | 0 | -- |
| KEWA | 9 | 11.1 | 4 | 25.0 | 14 | 28.6 | 2 | 0.0 | 1 | 0.0 |
| HOWA | 0 | -- | 0 | -- | 11 | 63.6 | 7 | 57.1 | 0 | -- |
| WEWA | 25 | 12.0 | 30 | 43.3 | 16 | 25.0 | 6 | 33.3 | 0 | -- |
| OVEN | 25 | 8.0 | 11 | 27.3 | 6 | 50.0 | 6 | 33.3 | 0 | -- |
| YBCH | 1 | 0.0 | 0 | -- | 3 | 0.0 | 0 | -- | 6 | 0.0 |
| NOCA | 13 | 30.8 | 14 | 7.1 | 12 | 8.3 | 4 | 25.0 | 0 | -- |
| INBU | 16 | 6.3 | 16 | 6.3 | 10 | 30.0 | 2 | 0.0 | 1 | 0.0 |
| EATO | 0 | -- | 0 | -- | 7 | 0.0 | 1 | 0.0 | 1 | 0.0 |
| SCTA | 2 | 50.0 | 2 | 100.0 | 2 | 100.0 | 0 | -- | 0 | -- |
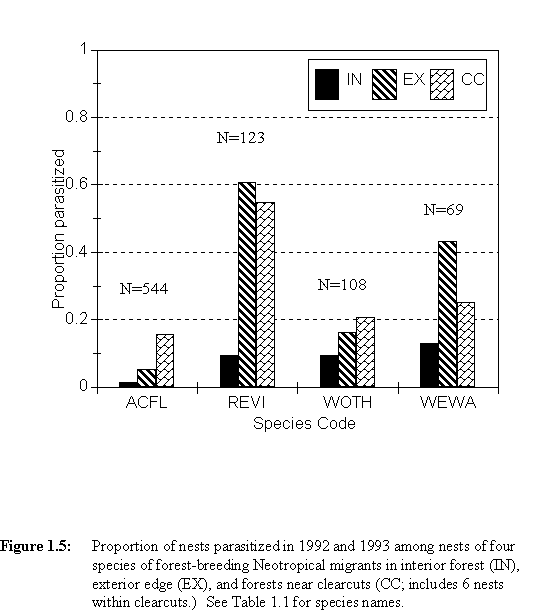 exact probability = 0.001) or interior forest (1.5%, p < 0.0005).
This species was parasitized more heavily in exterior than interior, although
the difference was not quite significant (p = 0.087). Red-eyed Vireos, however,
were parasitized as heavily in exterior edge (60.7%) as in forests near clearcuts
(54.8%, p = 0.793), and nests of this species were parasitized more heavily
in each of these edge contexts than in interior forest (9.4%, p < 0.0005
for both exterior and clearcuts). The results of these pairwise comparisons
were qualitatively similar when examined for each year separately.
exact probability = 0.001) or interior forest (1.5%, p < 0.0005).
This species was parasitized more heavily in exterior than interior, although
the difference was not quite significant (p = 0.087). Red-eyed Vireos, however,
were parasitized as heavily in exterior edge (60.7%) as in forests near clearcuts
(54.8%, p = 0.793), and nests of this species were parasitized more heavily
in each of these edge contexts than in interior forest (9.4%, p < 0.0005
for both exterior and clearcuts). The results of these pairwise comparisons
were qualitatively similar when examined for each year separately.
Table 1.3: Loglinear modeling of proportion of
parasitized nests with respect to landscape context and host species.
| Model | G | df | P |
| CONTEXT*SPECIES+PAR | 103.82 | 5 | 0.000 |
| CONTEXT*SPECIES+CONTEXT*PAR | 74.58 | 3 | 0.000 |
| CONTEXT*SPECIES+SPECIES*PAR | 61.51 | 4 | 0.000 |
| CONTEXT*SPECIES+CONTEXT*PAR+SPECIES*PAR | 5.40 | 2 | 0.067 |
| Effect of context | 29.24 | 2 | <0.001 |
| Effect of species | 42.31 | 1 | <0.001 |
Note: For Acadian Flycatcher and Red-eyed Vireo nests in continuous forest, at exterior edges, and in forests near clearcuts in 1992 and 1993, including three nests within clearcuts. The effect of context is tested by subtracting the log-likelihood ratio (G) and degrees of freedom for the model species X context + context X par from the respective statistics for the model species X context + par (Gokhale and Kullback 1978). The effect of species is tested in a similar manner. The degree of non-independence from the model species X context + context X par + species X par represents the effect of the interaction between species and context on parasitism.
The landscape pattern for the Worm-eating Warbler was similar to that seen for the Red-eyed Vireo, and this pattern was significant (G = 6.219, df = 2, p = 0.045, pooling data from 1992 and 1993). Pairwise comparisons of contexts, pooling both years, show that parasitism was higher in exterior edge (43.3%) than in the interior (13.0%) for this species (Fisher exact p = 0.033). The frequency of parasitism near clearcuts for this species appeared to be intermediate (25.0%) in degree, but was not significantly different from either of the other contexts (interior: p = 0.415; exterior: p = 0.338).
We monitored nests of several other species of ground-nesting warblers, but we do not have large samples for any one species. We therefore pooled data from all ground-nesting warblers in order to look for landscape patterns of parasitism in this suite of species. The species included in this sample are the Worm-eating Warbler, Ovenbird, Kentucky Warbler, Louisiana Waterthrush, Blue-winged Warbler, Black-and-white Warbler, and Common Yellowthroat. Proportion of parasitized nests did not differ between 1992 and 1993 for these species (Mantel-Haenszel chi-square = 0.001, p = 0.975). Proportion of parasitized nests for these species varied with landscape context (G = 13.130, df = 2, p = 0.001, pooling data from 1992 and 1993 and comparing interior, exterior, and forests near clearcuts). Parasitism was higher in the exterior (36.7%, p = 0.001) and clearcut edge (32.5%, p = 0.008) contexts than in the interior (10.0%) for these species. The level of parasitism did not differ significantly between exterior and forests near clearcuts (p = 0.823).
The overall level of parasitism in forest adjacent to forest openings in 1993 was high (13 of 78 nests, all species pooled, including nests in successional areas) relative to the parasitism level in interior forest for that year. Of 33 Acadian Flycatcher nests near forest openings, however, only one was parasitized.
We have a small sample of nests from within timbercuts. In 1992, 3 of 10 nests (30%) within cuts in HNF and 34 of 150 (22.7%) in forest near clearcuts were parasitized. In 1993, 2 of 25 nests (8%) within HNF clearcuts and 42 of 184 nests (22.8%) near clearcuts were parasitized. We also monitored 13 nests within and 2 nests in forest near small timbercuts in YSF in 1993, and none of these nests were parasitized. Pooling data from both types of cut and controlling for the effect of year, a test of independence of parasitism rate and habitat shows a non-significant trend toward higher parasitism in the forest near timbercuts than in the cuts themselves (Mantel-Haenszel chi-square = 3.416, p = 0.065). This difference reflects variation in species composition between forest and successional samples. For instance, 35.4% of the nests in successional areas (pooled over all sites and years) were of species which are only rarely parasitized or commonly reject cowbird eggs. Only 2.1% of the nests in mature forest were of such species.
The rate of parasitism in the old field site monitored in 1993 was low (2 of 38 nests, all species pooled, including 8 from rejectors and species rarely parasitized).
C. Parasitism in Relation to Edge Proximity
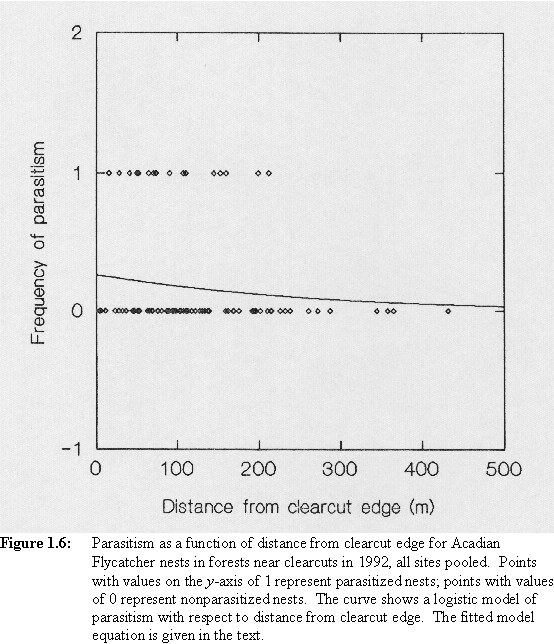
Logistic regression of parasitism of Acadian Flycatcher nests monitored in 1992 on distance to clearcut edge yielded the fitted model equation PAR = exp(-1.0424-0.0047*DISTANCE)/(1+exp(-1.0424-0.0047*DISTANCE)), where PAR = the probability that a nest is parasitized and DISTANCE = the distance in meters to the edge of the focal clearcut (Figure 1.6). The negative value for the coefficient of DISTANCE suggests that the incidence of parasitism decreased with distance to clearcut edge; however, this estimate was not significantly different from 0 (S.E.= 0.0039, p = 0.2272).
DISCUSSION
O'Conner and Faaborg
(1992) showed that cowbird densities increased in proximity to interior
and exterior edges in the Missouri Ozarks. The census data I present in
 this chapter do not strongly support the hypothesis that cowbird densities
are elevated near edges, but do suggest male cowbirds may occur more frequently
in close proximity (within 100 m) to forest openings, clearcuts, and utility
corridors than at points further from edges. Morning densities of cowbirds
were not higher near field edges than in interior forest. The numbers of
sites and the numbers of cowbirds detected at each site were low; also,
there appeared to be substantial variation among sites and years. These
factors make it difficult to detect any trends that may exist. These data
indicate that cowbirds are found throughout the forest in and around the
Pleasant Run Unit. The high degree of internal disturbance within these
forests may influence cowbirds to range throughout the landscape.
this chapter do not strongly support the hypothesis that cowbird densities
are elevated near edges, but do suggest male cowbirds may occur more frequently
in close proximity (within 100 m) to forest openings, clearcuts, and utility
corridors than at points further from edges. Morning densities of cowbirds
were not higher near field edges than in interior forest. The numbers of
sites and the numbers of cowbirds detected at each site were low; also,
there appeared to be substantial variation among sites and years. These
factors make it difficult to detect any trends that may exist. These data
indicate that cowbirds are found throughout the forest in and around the
Pleasant Run Unit. The high degree of internal disturbance within these
forests may influence cowbirds to range throughout the landscape.
Subjective observations suggest that cowbirds are more abundant near edges, although the data presented here do not strongly support this claim. The census plots did not include the disturbances themselves, but the forests surrounding them. As indicated by censuses conducted in 1992 to determine daily patterns of habitat use (unpublished data), cowbirds were present in clearcuts in high numbers in the early morning. I often see and hear them in clearcuts and forest openings, chasing and displaying from prominent perches. Additional censuses conducted at forest openings in 1995 (unpublished data) indicate that male cowbirds occur in higher numbers within forest openings than in nearby forest. These observations could be deceptive, as cowbirds may simply be more conspicuous in openings and thus more likely to be detected. Cowbirds are absent from these interior openings by the afternoon when they are feeding in nearby agricultural fields and lawns. This daily activity pattern has been documented for other populations of cowbirds through radio-telemetry (Rothstein et al. 1986, Thompson 1994, Thompson and Dijak 2000).
In more recent field seasons we have censused cowbirds and host species at sites where we have measured nesting success. These data will provide better estimates of cowbird and host densities and make it possible to relate these variables to landscape patterns and nesting success.
Levels of parasitism in our study area are low relative to other areas of the Midwest. Robinson and coworkers (Robinson 1992, Robinson et al. 2000) have documented extremely high parasitism in sites throughout Illinois. Faaborg and coworkers (Thompson et al. 2000) have found high levels of parasitism in fragmented sites in northern Missouri. Hoover and Brittingham (1993) analyzed data from Wood Thrush nests compiled by the Cornell Nest Record Program and reported frequencies of parasitism of over 40% for this species in Indiana and in the Midwest region as a whole. On the other hand, parasitism levels in the heavily forested Ozarks of Missouri are as low as 3% (Thompson et al. 2000). Parasitism pressure at regional scales is probably heavily influenced by landscape factors, such as percent forest cover and habitat heterogeneity. The landscape within which we work is heavily forested relative to areas in the Midwest where cowbird parasitism is high; this may explain the low levels of parasitism observed (Robinson et al. 1995).
In our study area, Red-eyed Vireos and ground-nesting warblers were more heavily parasitized than Acadian Flycatchers and Wood Thrushes. This contrasts with patterns in Illinois, where Wood Thrushes often suffer parasitism frequencies close to 100% and ground-nesting species usually exhibit lower levels of parasitism than understory and canopy nesting species (Robinson 1992, Robinson et al. 2000, Trine 2000). Such regional differences in host-specific parasitism frequencies may result from regional differences in the behavior of cowbirds, host species, or both. Cowbirds may actively follow certain host species. Which species are preferred in a given area may be a function of local community structure or species-specific levels of nest predation. Alternatively, variables such as host defense behavior or nest height may differ for a given species over regional scales, and this may affect the ability of cowbirds to find or the propensity for cowbirds to parasitize nests of that species. More information addressing the mechanisms of cowbird nest-searching and host defense behavior is needed.
These results indicate that levels of parasitism are lower in interior forest than in forests adjacent to exterior or interior edge. This provides support for the hypothesis that levels of parasitism are elevated in proximity to internal and external edge (e.g. Brittingham and Temple 1983). Such local landscape patterns of parasitism probably result to a large extent from the habitat selection behavior of cowbirds. Host species differ in landscape patterns of parasitism over local scales (Figure 1.5). For the Acadian Flycatcher, and perhaps the Wood Thrush, parasitism was higher near clearcuts than in the interior forest or exterior edge ravines. For the Red-eyed Vireo, and perhaps ground-nesting warblers, parasitism was as high or higher in the exterior as in forests near clearcuts.
Levels and within-landscape patterns of parasitism were quite similar in 1992 and 1993, but data from 1994 (unpublished) indicate that annual variation may be important. In the two HNF clearcut sites that were monitored in that year, only 2 of 50 Acadian Flycatcher nests were parasitized; whereas 6 of 66 were parasitized in interior forest. This may also reflect variation among sites within a landscape context. Data from the five clearcuts were pooled in this analysis; however, these sites vary in measured levels of parasitism and in landscape variables that may be related to parasitism intensity (P. Doran et al., unpublished data).
Initial results suggest that levels of parasitism may be lower within successional areas than in surrounding forest. The data are difficult to interpret, however, because sampling effort has varied between years. A larger sample of nests from small timbercuts and adjacent forest in YSF in 1994 showed this same pattern (0 of 26 nests within cuts and 27 of 170 in forest near cuts parasitized; T. Ford, unpublished data). This pattern may result from differences in host species composition between these two habitats. A large proportion of the sample of nests from timbercuts consisted of species that are only rarely parasitized and species that reject cowbird eggs. Early successional species may be more likely to have evolved defenses against parasitism, such as egg rejection (Mayfield 1965). Also, cowbirds may prefer to parasitize forest species. Several recent studies have found parasitism rates to be low in successional areas (Robinson et al. 2000).
Data from 1992 suggest that parasitism of Acadian Flycatcher nests declines with distance from clearcut edge, although the result is not statistically significant. I have information that will make it possible to calculate distances from nests monitored in other years and other edge contexts, which should enable the detection of trends (if they exist) of the magnitude suggested by the model. Data from forest openings suggest that parasitism decreases with distance to forest opening edge (P. Doran, unpublished).
Comparison of these results with those of other studies shows that parasitism levels vary among species and over both large and small spatial scales. Furthermore, spatial patterns of parasitism at large and small scales vary with species. This may result from variation in the behavior of cowbirds and/or hosts. Because cowbird females have large breeding ranges and are often territorial (Rothstein et al. 1986, Thompson 1994), it is conceivable that the behavior of individual females could greatly influence patterns of parasitism at local scales.
These results have important implications for both research and management. In terms of research, it is clear that the results of field studies performed in one location cannot be used to predict confidently landscape patterns of parasitism for individual species in other areas. Coordinated intensive research efforts over large spatial scales will be necessary to understand the factors that determine the frequency and pattern of parasitism.
In terms of management, it is evident that disturbance and fragmentation at all spatial scales can increase levels of parasitism in forest-breeding Neotropical migrant birds (NTMB). Parasitism levels are elevated in proximity to both internal and external edge. It is not yet clear how parasitism pressure affects the population dynamics of NTMB. When combined with other deleterious effects of forest fragmentation such as reduced habitat availability and increased nest predation, brood parasitism may seriously threaten NTMB populations. If we are to preserve viable populations of forest-breeding NTMB, it may be necessary to maintain a network of large continuous tracts of forest and to minimize the amount of internal edge. The forests of south-central Indiana may constitute an important source for populations of NTMB. Management activities presently occurring in state and national forests, such as timber harvest and the creation and maintenance of forest openings, increase the area of internal edge habitat. Such habitat alteration may reduce nesting success and thus detract from this landscape's value as a source for populations of Neotropical migrant birds.